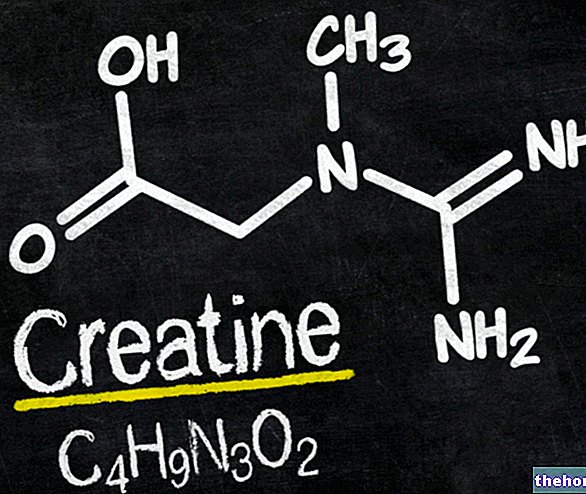
" prvá časť
Enzým, ktorý katalyzuje štvrtú fázu Krebsovho cyklu, je a-keto glutarát dehydrogenáza; tento enzým je komplex enzýmov veľmi podobný pyruvátdehydrogenáze. Oba sú zložené z 48-60 proteínov, v ktorých sú rozpoznané tri rôzne enzymatické aktivity a majú tiež rovnaké enzymatické kofaktory; sú veľmi podobné enzýmy, pretože pôsobia na podobné substráty: pyruvát aj l "α-keto glutarát, sú α- keto kyseliny. Mechanizmus účinku týchto dvoch enzymatických komplexov je rovnaký.
Útok tiamín pyrofosfátu na karbonyl (C = O)α-ketoglutarát, vedie k jeho dekarboxylácii a vzniká karboxyhydroxypropylový derivát. S následným prenosom na lipoamid prebieha vnútorný redoxný proces, z ktorého sa získa lipoamid karboxyderivát alebo sukcinyl lipoamid.
Sukcinyl lipoamid potom reaguje s koenzýmom A za vzniku sukcinyl koenzýmu A (ktorý pokračuje v Krebsovom cykle) a redukovaný lipoamid, ktorý sa reoxiduje FAD: vytvorený FADH2 sa znova oxiduje NAD + a získa sa NADH. V tejto fáze preto došlo k druhej eliminácii uhlíka z uhlíkatého skeletu vo forme oxidu uhličitého.
Acylová skupina viazaná na koenzým A je v aktivovanej forme, to znamená, že má vysoký obsah energie: je teda možné využiť energiu sukcinyl koenzýmu A.

V piatej fáze Krebsovho cyklu je sukcinyl koenzým A vystavený pôsobeniu sukcinyltiokináza; o jeho spôsobe pôsobenia boli urobené dve hypotézy: opíšeme iba jeden z týchto dvoch, pretože je najviac akreditovaný. Podľa tejto hypotézy je sukcinyl koenzým A napadnutý dusíkom histidínu (Hys) enzýmu: koenzým A sa uvoľňuje a adukt odvodený od histidínu sa tvorí ako medziprodukt, tj. Sukcinyl-enzým (alebo sukcinyl-Hys Na tento medziprodukt pôsobí ortofosfát, čo vedie k uvoľňovaniu sukcinátu a tvorbe fosfoenzýmu. Fosfoenzým napadnutý guanozíndifosfátom (GDP) produkuje guasnosíntrifosfát (GTP) a enzým sa uvoľňuje. Z energetického hľadiska GTP = ATP: väzba, ktorá dodáva energiu, je v oboch druhoch rovnaká (jedná sa o anhydridovú väzbu medzi fosforylom Β a fosforylom γ). V niektorých prípadoch sa GTP používa ako materiál s vysokým obsahom energie, ale zvyčajne sa GTP premieňa na ATP pôsobením enzýmu. nukleozid difosfokináza (NDPK); je enzým nachádzajúci sa v bunkách a katalyzuje nasledujúcu reakciu:
N1TP + N2DP → N1DP + N2TP
Generický NiTP ® nukleozid trifosfát
Generický nukleozid difosfát NiDP®
Je to reverzibilná reakcia; v našom prípade sa stane:
GTP + ADP → GDP + ATP
preto môže postupovať doprava alebo doľava aj pri malých odchýlkach v koncentráciách reagencií.
Ak krebsov cyklus prebieha takou rýchlosťou, ktorá vedie k produkcii ATP vyššej, ako je energetická potreba, je ADP málo dostupná, zatiaľ čo existuje veľa ATP: reakcia katalyzovaná nukleozidovou difosfokinázou je, potom, namierené doľava (GTP sa hromadí, ak nukleoziddifosfokináza nemá dostatočný substrát, t.j. ADP). GTP je preto signálom dostupnosti energie, a preto spomaľuje Krebsov cyklus.
Šiesta etapa krebsovho cyklu vedie k vzniku fumarátu pôsobením sukcinát dehydrogenáza; tento enzým dáva stereošpecifickú reakciu, pretože vždy vzniká nenasýtený (je to alkén) trans, tj. fumarát (zatiaľ čo cis izomér je maleát). Sukcinát dehydrogenáza sa nachádza na vnútornej mitochondriálnej membráne, zatiaľ čo všetky ostatné enzýmy Krebsovho cyklu sú rozptýlené v mitochondriách.
Sukcinát dehydrogenáza má ako kofaktor FAD; je inhibovaný oxaloacetátom (inhibícia spätnej väzby), zatiaľ čo má sukcinát a fumarát ako svoj pozitívny modulátor (aktivátor). jeho aktivátor. Pokúsme sa pochopiť, prečo skokom do konečnej fázy Krebsovho cyklu. Krebsov cyklus vyžaduje energiu, takže jedinou možnosťou, ako získať oxaloacetát od pacienta, je to, že jeho koncentrácia je veľmi vysoká: malát je jedným z metabolitov s najvyššou koncentráciou v bunkách. Reakcia, ktorá prevádza malát na oxaloacetát, je tiež obľúbená skutočnosť, že koncentrácia oxaloacetátu je udržiavaná na nízkej úrovni pôsobením citrát syntázy. Reakcia katalyzovaná sukcinátdehydrogenázou je potom samokŕmna reakcia a toto je jediný spôsob, ako uskutočniť transformáciu malátu na oxaloacetát.
Koncentrácia mitochondriálneho malátu musí byť kompatibilná s koncentráciou cytoplazmatického malátu: iba vtedy, keď je koncentrácia mitochondriálneho malátu taká vysoká, že zaručuje premenu malátu na oxaloacetát (v Krebsovom cykle), potom sa malát môže použiť aj v inými spôsobmi (ktoré sú cytoplazmatické): v cytoplazme môže byť malát premenený na oxaloacetát, z ktorého je možné získať aspartát pôsobením GOT (jedná sa o transaminázu) alebo glukózu glukoneogenézou.
Vraciame sa do siedmej fázy Krebsovho cyklu, ktorá je katalyzovaná enzýmom fumarasi: voda sa pridá stereošpecifickým spôsobom na výrobu L-malátu.
V poslednej fáze Krebsovho cyklu, o ktorej sme už hovorili, ide o akciu malátdehydrogenáza. Tento enzým používa na svoje katalytické pôsobenie molekulu NAD +.
Tým sme uzavreli popis rôznych fáz Krebsovho cyklu.
Krebsov cyklus je úplne reverzibilný.
Na zvýšenie rýchlosti Krebsovho cyklu je možné zvýšiť koncentráciu metabolitov prítomných v tomto cykle; jedna zo stratégií na zvýšenie rýchlosti Krebsovho cyklu spočíva v premene časti pyruvátu, ktorý vstupuje do mitochondrií, na oxaloacetát (pôsobením pyruvátkarboxylázy) a nie v jeho transformácii na acetylkoenzým A: čím sa zvyšuje koncentrácia oxaloacetátu, ktorá je metabolitom Krebsovho cyklu, a preto zvyšuje rýchlosť celého cyklu.
V Krebsovom cykle sa tri NAD + konvertujú na tri NADH a jeden FAD na FADH2 a navyše sa získa GTP: usmernením redukčného výkonu získaného z Krebsovho cyklu sa produkuje ďalší ATP; v dýchacom reťazci sa redukčná sila prenáša z NADH a FADH2 na kyslík: tento prenos je spôsobený sériou enzýmov umiestnených na mitochondriálnej membráne, ktoré svojim pôsobením vedú k produkcii ATP.
Procesy dýchacieho reťazca sú exergonické procesy a uvoľnená energia sa používa na produkciu ATP; účelom bunky je využiť exergonické procesy na uskutočnenie syntézy ATP. Pre každú molekulu NADH, ktorá vstupuje do dýchacieho reťazca, sa získa 2,5 molekuly ATP a pre každý FADH2 1,5 molekuly ATP; táto rozmanitosť je spôsobená skutočnosťou, že FADH2 vstupuje do dýchacieho reťazca na nižšej úrovni ako NADH.
Pri redukčnej sile aeróbneho metabolizmu sa získa 30-32 ATP (219-233 kcal / mol) s účinnosťou asi 33% (účinnosť anaeróbneho metabolizmu je asi 2%).