Upravil Dr. Giovanni Chetta
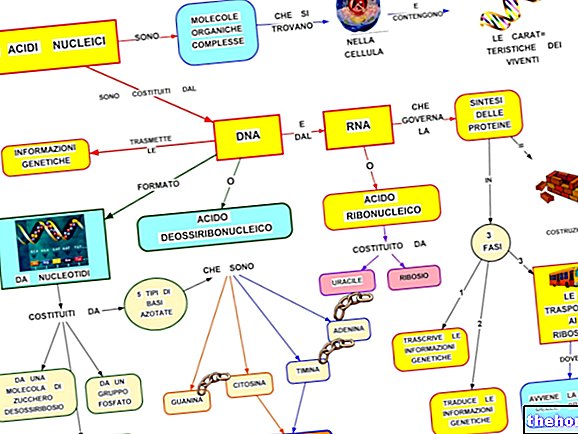
ECM sa všeobecne opisuje ako zložený z niekoľkých veľkých tried biomolekúl:
- Štrukturálne proteíny (kolagén a elastín)
- Špeciálne proteíny (fibrilín, fibronektín, laminín atď.)
- Proteoglykány (agregáty, syndekany) a glusaminoglykány (hyaluronany, chondroitínsulfáty, heparánsulfáty atď.)
Štrukturálne proteíny
Kolagény tvoria najpočetnejšie zastúpenú skupinu glykoproteínov v živočíšnej ríši. Sú to najprítomnejšie proteíny v extracelulárnej matrix (ale nie najdôležitejšie) a sú základnými zložkami vlastných spojivových tkanív (chrupavka, kosť, fascia, šľachy, väzy).
Existuje najmenej 16 rôznych typov kolagénu, z ktorých typy I, II a III sú najbežnejšie prítomné na úrovni typických fibríl (typ IV tvorí druh retikula, ktoré predstavuje hlavnú zložku bazálnych lamiel).
Kolagény sú väčšinou syntetizované fibroblastmi, ale syntetizovať ich môžu aj epiteliálne bunky.
Kolagénové vlákna nepretržite interagujú s obrovským množstvom ďalších molekúl extracelulárnej matrice, čo predstavuje biologické kontinuum zásadné pre život bunky. Príslušné kolagény vo vláknach zohrávajú vedúcu úlohu pri vytváraní a udržiavaní štruktúr schopných odolávať napínacím silám, je takmer neelastický (glukozamidy môžu vykonávať účinok odolnosti voči stlačeniu). Kolagén sa nejakým spôsobom produkuje a metabolizuje v závislosti od mechanického zaťaženia a jeho viskoelastické vlastnosti, ako uvidíme v odseku „Viskoelasticita fascia “, veľký vplyv na držanie tela človeka. Ako ďalšia demonštrácia schopnosti kolagénu sa meniť podľa vplyvov prostredia, za predpokladu, že napr. variabilným stupňom tuhosti, pružnosti a odolnosti, existujú kolagény, definované pojmom FACIT (kolagén asociovaný s vláknami s prerušenými trojitými závitovkami), schopné fungovať funkčne ako proteoglykány (popísané v odseku „Glukozaminoglykány a proteoglykány“).
Kolagénové vlákna vďaka svojmu povlaku z PG / GAG (proteoglykány / glukozaminoglykány) majú vlastnosti biosenzorov a biovodičov: relatívne elektrické náboje majú za následok väčšiu schopnosť viazať vodu a vymieňať ióny, a tým aj väčšiu elektrickú kapacitu.
Vieme, že akákoľvek mechanická sila schopná generovať štrukturálnu deformáciu namáha medzimolekulárne väzby a vytvára mierny elektrický tok, tj. piezoelektrický prúd (Athenstaedt, 1969). V takýchto prípadoch kolagénové vlákna distribuujú pozitívne náboje na ich konvexný povrch a negatívne na konkávnom povrchu, čím sa transformujú na polovodiče (umožňujú tok elektrónov na ich jednosmernom povrchu). Pretože piezoelektrická energia (ako aj pyroelektrická energia generovaná tepelnými pnutiami) je cirkulujúcimi iónmi neutralizovaná vo veľmi krátkom čase (približne 10-7-10-9 sekúnd), usporiadanie PG / GAG na signáli je rozhodujúci pre šírenie signálu.povrch fibrilov, ktoré pôsobia ako "opakovače" elektrického impulzu. Najmä pozdĺžna periodicita cca. 64 nm (čo sa pod optickým mikroskopom javí ako pruh) umožňuje rýchlosť šírenia impulzu približne 64 m / s (čo zodpovedá rýchlosti vedenia rýchlych nervových vlákien) - Rengling, 2001. Silný dipolárny moment kolagénových fibríl a ich rezonančná kapacita (vlastnosť spoločná pre všetky peptidové štruktúry), ako aj nízka dielektrická konštanta MEC uľahčujú prenos elektromagnetických signálov. Preto trojrozmerná a všadeprítomná kolagénová sieť má tiež zvláštnu vlastnosť vedenia bioelektrických signálov v tri rozmery priestoru, založené na relatívnom usporiadaní medzi kolagénovými vláknami a bunkami, v aferentnom smere (od ECM k bunkám) alebo naopak, eferentnom.
To všetko predstavuje komunikačný systém MEC-buniek v reálnom čase a takéto elektromagnetické biosignály môžu viesť k dôležitým biochemickým zmenám, napríklad v kosti osteoklasty nedokážu „stráviť“ piezoelektricky nabitú kosť (Oschman, 2000).
Na záver je potrebné zdôrazniť, že bunka neprekvapuje, že produkuje nepretržite a so značným výdajom energie (približne 70%) materiál, ktorý musí byť nevyhnutne vylúčený, a to väčšinou prostredníctvom výhradného skladovania protokolagénu (biologického prekurzora kolagénu) v špecifických vezikuly (Albergati, 2004).
Prevažná väčšina tkanív stavovcov vyžaduje súčasnú prítomnosť dvoch životne dôležitých vlastností: pevnosti a pružnosti. Skutočná sieť elastických vlákien umiestnená vo vnútri ECM týchto tkanív umožňuje návrat do pôvodných podmienok po silných trakciách. Elastické vlákna sú schopné zvýšiť roztiahnuteľnosť orgánu alebo jeho časti najmenej päťkrát. Dlhé, nepružné kolagénové vlákna sú rozptýlené medzi elastickými vláknami s presnou úlohou obmedziť „nadmernú deformáciu v dôsledku ťahu tkanív. L“elastín predstavuje hlavnú zložku elastických vlákien. Jedná sa o extrémne hydrofóbny proteín s dĺžkou približne 750 aminokyselín, pretože kolagén je bohatý na prolín a glycín, ale na rozdiel od kolagénu nie je glykovaný a obsahuje veľa zvyškov hydroxyprolínu, a nie hydroxylisin. Elastín sa javí ako skutočná biochemická sieť nepravidelne trojrozmerného tvaru, ktorá je zložená z vlákien a lamiel, ktoré prestupujú ECM všetkých spojivových tkanív. Nachádza sa v obzvlášť hojnom množstve v cievach s elastickými vlastnosťami (je to proteín ECM prítomný v tepnách a predstavuje viac ako 50% celkovej suchej hmotnosti aorty), vo väzivách, v pľúcach a v koži. V dermis, na rozdiel od toho, čo sa deje s kolagénom, hustota a objem elastínu majú tendenciu sa časom zvyšovať, ale starý elastín sa zdá byť opuchnutý, takmer opuchnutý, často s fragmentárnym vzhľadom a s redukciou zložky. „Amorfný“ (Pasquali Rochetti a kol., 2004). Bunky hladkého svalstva a fibroblasty sú hlavnými producentmi jeho prekurzora, tropoelastínu, vylučovaného v extracelulárnych priestoroch.
Ďalšie články na tému „Kolagén a elastín, kolagénové vlákna v extracelulárnej matrix“
- Extracelulárnej matrix
- Fibronektín, glukozaminoglykány a proteoglykány
- Význam extracelulárnej matrice v bunkových rovnováhach
- Zmeny extracelulárnej matrice a patológie
- Spojivové tkanivo a extracelulárna matrica
- Hlboká fascia - spojivové tkanivo
- Fasciálne mechanoreceptory a myofibroblasty
- Biomechanika hlbokej fascie
- Držanie tela a dynamická rovnováha
- Tensegrity a špirálové pohyby
- Pohyb dolných končatín a tela
- Podpora záveru a stomatognatický aparát
- Klinické prípady, posturálne zmeny
- Klinické prípady, držanie tela
- Posturálne hodnotenie - klinický prípad
- Bibliografia - Od extracelulárnej matrice k držaniu tela. Je spojivový systém náš skutočný Deus ex machina?