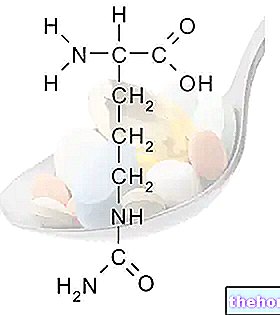
„Prvé kroky glykolýzy
Prvým enzýmom použitým v druhej fáze glykolýzy je glyceraldehyd 3-fosfát dehydrogenáza; dehydrogenázy sú enzýmy, ktoré katalyzujú prenos redukčnej sily z redukujúcej molekuly, ktorá oxiduje na inú molekulu, ktorá je redukovaná (redoxná reakcia). Substrátmi tohto enzýmu sú NAD (nikotidamidadenin dinukleotid) a FAD (flavinadenin dinukleotid).
V tomto kroku dehydrogenáza katalyzuje premenu glyceraldehyd-3-fosfátu na 1,3-bisfosfoglycerát: na tom istom katalytickom mieste sa aldehydová skupina oxiduje na karboxyl s následnou redukciou NAD + na NADH a následne je karboxylová skupina schopná za vzniku anhydridovej väzby s ortofosfátom. Prvý proces je veľmi exergonický (uvoľňuje energiu), zatiaľ čo druhý je veľmi exergonický (vyžaduje energiu); keby neexistovalo žiadne katalytické miesto, globálna reakcia by neprebehla: prvá reakcia by nastala s uvoľnením energie, ktorá by bola dispergovaná ako teplo a ktorá by preto nebola použiteľná na vytvorenie anhydridovej väzby.

Po vytvorení 1,3-bisfosfoglycerátu enzým obnoví svoju pôvodnú štruktúru a je pripravený pôsobiť na nový substrát.

Potom príde fosfoglycerátkináza ktorý umožňuje prenos fosforylu z 1,3-bisfosfoglycerátu na molekulu ADP; získali sme ATP (jeden ATP pre každú molekulu glyceraldehyd-3-fosfátu, teda dva ATP pre každú počiatočnú molekulu glukózy), ktorý kompenzuje energetický výdaj prvá fáza glykolýzy.
Arseničitanový anión (AsO43-) ovplyvňuje glykolytickú dráhu, pretože môže nahradiť fosfát v prvej reakcii druhej fázy glykolýzy, čím sa získa 1-arzenio-3-fosfoglycerát, ktorý je vysoko nestabilný a akonáhle je bez katalyzátora miesto, hydrolyzuje a uvoľňuje „arzenát vracajúci sa do obehu. Arzenát preto napodobňuje pôsobenie fosfátu a vstupuje do katalytického miesta: v prítomnosti arzenátu reakcia, ktorá produkuje ATP (od 1,3-bisfosfoglycerátu po 3-fosfoglycerát), neprebieha, pretože 3-fosfát glyceraldehyd sa premieňa priamo na 3-fosfoglycerát; bez dostupnosti ATP bunky odumierajú (otrava kyselinou arzénovou).
V tretej reakcii oxidačnej fázy sa 3-fosfoglycerát premení na 2-fosfoglycerát pôsobením fosfoglycerát mutáza; reakcia zahŕňa 2,3-bisfosfoglycerátový medziprodukt.

V ďalšom kroku zasiahne enzým enolase ktorý je schopný katalyzovať elimináciu molekuly vody z uhlíkatého skeletu 2-fosfoglycerátu za získania pyrivizovaného fosfoenolu (PEP);

PEP má vysoký potenciál prenosu fosforylu: prenáša sa pôsobením enzýmu pyruvátkináza, fosforyl na ADP za vzniku ATP, v piatom kroku druhej fázy, získanie pyruvátu.

2-fosfoglycerát a 3-fosfoglycerát majú nízku prenosovú silu fosforylu, a preto sa na získanie ATP z týchto molekúl 3-fosfoglycerát počas glykolýzy premieňa na 2-fosfoglycerát, pretože sa z neho získava. PEP, ktorý je druhy s vysokým potenciálom prenosu.
Pred pokračovaním otvorme zátvorku na 2,3-bisfosfoglyceráte; ten posledný je prítomný vo všetkých bunkách, v ktorých prebieha glykolýza vo veľmi nízkej koncentrácii (je to medziprodukt tretej reakcie druhej fázy glykolýzy). V erytrocytoch má naopak 2,3-bisfosfoglycerát stacionárnu koncentráciu 4-5 mM (maximálna koncentrácia), pretože majú enzymatické dedičstvo, ktoré má za úlohu ho produkovať; v erytrocytoch dochádza k odchýlke od glykolýzy za vzniku 2,3-bisfosfoglycerátu: 1,3-bisfosfoglycerát sa prevádza na 2,3-bisfosfoglycerát bisfosfoglycerát mutáza (erytrocyt) a 2,3-bisfosfoglycerát, pôsobením bisfosfoglycerát fosfatáza (erytrocyt) sa stáva 3-fosfoglycerátom. Potom sa v erytrocytoch časť 1,3-bisfosfoglycerátu získaného z glykolýzy premení na 2,3-bisfosfoglycerát, ktorý sa potom vráti na glykolytickú dráhu ako 3-fosfoglycerát; pri tom je tretím krokom oxidačnej fázy glykolýza, z ktorej sa získava ATP. Množstvo strateného ATP je cena, ktorú je erytrocyt ochotný zaplatiť za udržanie koncentrácie 2,3-bisfosfoglycerátu, ktorú tieto bunky potrebujú, pretože ovplyvňuje schopnosť „hemoglobínu viazať“ kyslík.
Videli sme, že v prvej reakcii druhej fázy glykolýzy je NAD + redukovaný na NADH, ale je nevyhnutné, aby sa po získaní pyruvátu NADH znova premenil na NAD +: k tomu dochádza pri mliečnom kvasení (získava sa laktát) alebo alkoholovým kvasením (do hry vstupuje pyruvát dekarboxyláza, ktorá dekarboxyluje pyruvát, a dehydrogenáza, ktorá tvorí etanol); fermentácie nezahŕňajú kyslík (anaeróby).
V dôsledku mliečneho kvasenia sa kyselina mliečna, ak nie je dostatočne zneškodnená, hromadí vo svaloch a uvoľňuje H +, spôsobuje mimovoľné sťahovanie svalov, a preto kŕče; sval v silnom strese môže tiež dosiahnuť minimálne pH 6,8.
Prostredníctvom Coriho cyklu sa časť únavy svalu prenáša na pečeň, keď je sval preťažený. Predpokladajme, že sval pracuje bez prísunu kyslíka (nesprávny predpoklad): ak sval pracuje striedmo, ATP potrebný na kontrakciu poskytuje výlučne glykolýza. Ak sa aktivita svalu zvýši a je potrebný ďalší ATP, zrýchlite aeróbny metabolizmus, laktát, ktorý sa takto zlikviduje, na glukózu. V skutočnosti sval využíva aeróbny metabolizmus: ak je k dispozícii kyslík, sval využíva predovšetkým ATP poskytovaný aeróbnym metabolizmom, a keď už nie je k dispozícii žiadny kyslík, anaeróbny metabolizmus sa zrýchľuje prostredníctvom Coriho cyklu. Tento cyklus predpokladá, že laktát sa prenáša zo svalu do pečene, kde výdajom energie produkuje viac glukózy, ktorá sa vracia do svalu. Prostredníctvom tohto cyklu časť ATP spotrebovaného vo svale dodáva pečeň, ktorá, prostredníctvom procesu glukoneogenézy je schopný produkovať glukózu, ktorú môže sval použiť na získanie ATP.
Doteraz popísaný metabolizmus glukózy nezahŕňa kyslík, ale aeróbny metabolizmus glukózy umožňuje získať 17-18-krát vyššie množstvo ATP, ako je množstvo získané pomocou glykolytickej dráhy, keď má bunka možnosť vybrať si medzi aeróbnym a ed. anaerób, uprednostňuje to prvé.
Pri aeróbnom metabolizme vstupuje pyruvát do mitochondrií, kde prechádza transformáciami a nakoniec sa získa oxid uhličitý a voda; týmto spôsobom sa získa 34 molekúl ATP pre každú molekulu degradovanej glukózy.