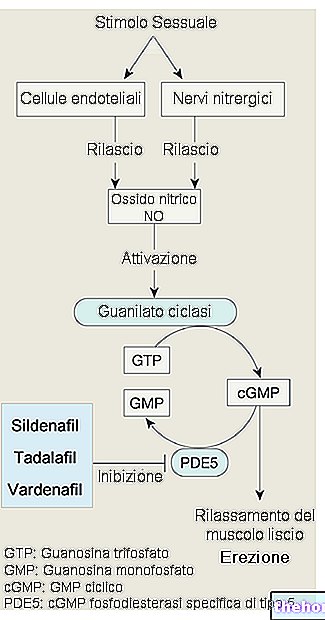
Ak bunka pracuje v anaeróbnych podmienkach, vyrába energiu premenou glukózy na laktát a prostredníctvom Coriho cyklu ju zneškodňuje; ak je k dispozícii kyslík (teda v kľudových podmienkach), viac ako 90% glukózy sa spotrebuje aeróbne a iba zvyšných 10%, anaeróbne. Keď existuje potreba väčšieho množstva ATP, ako je schopná poskytnúť aeróbna dráha (napríklad keď sú svaly pod stresom), potom je dodatočná zásoba zabezpečená anaeróbnym metabolizmom (nachádzame sa v podmienkach nedostatku kyslíka: dýchavičnosť, únava pri dýchaní a pod.): je potrebné urýchliť tento metabolizmus premenou laktátu (ktorý sa získava z glykolýzy) na glukózu glukoneogenézou.
V mitochondriách sa rozvíja aeróbny metabolizmus.
Prvým enzýmom, s ktorým sa stretávame v aeróbnom metabolizme, je pyruvátdehydrogenáza; Je presnejšie povedať, že pyruvát dehydrogenáza je skôr enzýmový komplex ako enzým, pretože ide o agregát 48-60 proteínových jednotiek s tromi katalytickými miestami pôsobiacimi za sebou.
Pyruvátdehydrogenáza katalyzuje nasledujúcu reakciu (redox):
Pyruvát + NAD + + CoA-SH → Acetyl CoA + NADH + H + + CO2
CoA-SH je koenzým A: je to derivát kyseliny pantoténovej; acetylkoenzým A je tioester. Ide o redoxný proces, pretože prvý uhlík pyruvátu prechádza z oxidačného čísla tri na oxidačné číslo štyri (oxiduje) a druhý uhlík pyruvátu prechádza z oxidačného čísla dva na oxidačné číslo tri (oxiduje). Potom sa pyruvát zoxiduje (stratí celkom dva elektróny) a NAD sa zníži.
Ako bolo uvedené, pyruvátdehydrogenáza má tri typy enzymatickej aktivity, z ktorých každý je podporovaný vlastným katalytickým kofaktorom:
- tiamín pyrofosfát (je to derivát vitamínu B1); je aktívny v deprotonovanej forme: vytvorí sa karbanion.
- lipoamid (je to derivát kyseliny lipoovej); obsahuje veľmi reaktívny disulfidový mostík.
- flavin adenin dinukleotid (je to derivát vitamínu B2); je to nukleotid s redoxnými vlastnosťami: jeho redoxné centrum je tvorené flavínom.
V eukaryotických bunkách dochádza k aeróbnemu metabolizmu v špecializovaných organelách bunky, ktorými sú mitochondrie; v baktériách dochádza k metabolizmu glukózy a iných druhov v bunke, ale neexistujú žiadne špecializované organely.
Keď pyruvát vstúpi do mitochondrií, je vystavený „pôsobeniu pyruvátkarboxylázy, ak je potrebné vykonať glukoneogenézu (na rekonštrukciu východiskového materiálu), alebo môže byť podrobený pyruvátdehydrogenáze, ak je to potrebné na výrobu energie: „Acetylkoenzým A, ktorý je tvorený aeróbnym metabolizmom, stimuluje pôsobenie pyruvátkarboxylázy, preto podporuje glukoneogenézu a znižuje pôsobenie pyruvátdehydrogenázy.
Pozrime sa teraz, ako pyruvátdehydrogenáza funguje; v prvom rade dochádza k dekarboxylácii pyruvátu pôsobením tiamín pyrofosfátu.
Kyslé prostredie môže inhibovať aeróbny metabolizmus, pretože je aktívna aniónová forma tiamín pyrofosfátu, ktorá by bola protonizovaná pri kyslom pH a nedošlo by k dekarboxylácii.
Dekarboxylácia je náročná reakcia, pretože je potrebné prerušiť väzbu uhlík-uhlík; v tomto prípade je reakcia termodynamicky zvýhodnená skutočnosťou, že reakčný medziprodukt (hydroxyetyl-tiamín pyrofosfát) dáva rezonanciu (p-elektróny molekuly sú delokalizované): hydroxyetyl-tiamín pyrofosfát existuje v troch možných formách (rezonancia) a vďaka tomu je celkom stabilný. Hydroxyetyl-tiamínpyrofosfát v aniónovej forme navyše prežíva dostatočne dlho, aby mohol interagovať s disulfidovým mostíkom lipoamidu (druhý katalytický kofaktor pyruvátdehydrogenázy); disulfidový mostík je oscilačné rameno (nachádza sa na koniec dlhého flexibilného reťazca) a môže sa pohybovať v enzýmovom komplexe z jedného katalytického miesta na druhé.
Potom sa lipoamid cez disulfidový mostík viaže na hydroxyetyl-tiamínpyrofosfát: získa sa acetyllipoamid. Toto je prvá fáza transacetylačnej reakcie katalyzovanej prvým enzýmom komplexu pyruvátdehydrogenázy; v tejto fáze sa prerušila väzba medzi hydroxylová skupina a tiamín pyrofosfát, ktoré sa vrátili do pôvodnej formy: prebiehala redoxná reakcia, pri ktorej disulfidový mostík pôsobil ako oxidačné činidlo (dva atómy síry sa redukovali) smerom k hydroxylovej skupine, ktorú oxidoval na acetyl.
Po tejto fáze sa oscilačné rameno lipoamidu pohybuje a približuje sa k druhému enzýmu pyruvátdehydrogenázy, ktorý vykonáva skutočnú transacetylázovú aktivitu tým, že nesie so sebou acetylovú skupinu: prebieha druhá fáza transacetylačnej reakcie katalyzovaná druhým enzýmom; týmto spôsobom sme získali acetylkoenzým A. Teraz je potrebné obnoviť lipoamid, ktorý je v redukovanej forme: zasahuje tretí enzým pyruvátdehydrogenázy, ktorý redoxuje lipoamid a prenáša svoje elektródy na FAD, ktorý je redukovaný na FADH2. FAD / FADH2 môže fungovať ako redoxný pár v dvoch odlišných monoelektronických stupňoch alebo v jednom bielektronickom stupni.
FADH2 okamžite odovzdá svoje elektróny NAD + a získa FAD a NADH + H +.
Acetylkoenzým A, získaný podľa opisu, je východiskovým produktom pre Krebsov cyklus (alebo cyklus trikarboxylových kyselín).